Team:Oxford/Test/Modeling
Modelling
Introduction
Mathematical modelling plays a crucial role in Synthetic Biology by acting as a link between the conception and the physical realisation of a biological circuit. Our modelling team has focussed on building a better picture of the project to evaluate the effectiveness of initial designs, as well as to provide insight into how the system can (or must) be improved. To do this we have split our modelling efforts into three main sections: Characterising our Cells, Interaction with the Environment, and Interaction with the Biofilm. By combining the information gathered in each of these section we hope to ultimately answer the question: Is our system feasible? If not, where should the design be altered?
To help readers of all kinds and specialisations understand this page we have produced guides for all the modelling techniques used in this section which are available on the Modelling Tutorial page and will be linked to when relevant on this page.

Characterising Our Cells
In this section we look at our cells in isolation in order to assess their functionality and answer important questions such as “How long does it take to produce a certain concentration of product?”, and “What are the main limiting rates/concentrations?”. The first will help us assess the feasibility of our project, ie are our cells too slow? The second will aid us in further optimising our design.
Arabinose Induced
We have decided to use an arabinose induced promoter for the expression of a number of our proteins. This promoter can be modelled as the following chemical system:
\[[Arab:AraC]\overset{K}{\rightarrow}mRNA\overset{\alpha{\rightarrow}P\] \[mRNA\overset{\gamma_{1}}{\rightarrow}\phi\quad P\overset{\gamma_{2}}{\rightarrow}\phi\]You can find out more information about how this promoter works here. For this system we will assume that AraC is always in large concentration and that it's binding to arabinose happens on a faster time scale to transcription. Therefore, we do not need to consider the individual concentrations of arabinose and AraC, instead we just need to include the concentration of the complex [Arab:AraC].
Using this approximation, we arrive at the equations:
\[\dfrac{d[mRNA]}{dt}=K_{max}\dfrac{[Arab:AraC]^{n}}{K_{half}^{n}+[Arab:AraC]^{n}}-\gamma_{1}[mRNA]\] \[\dfrac{d\left[P\right]}{dt}=\alpha\left[mRNA\right]-\gamma_{2}\left[P\right]\]Where we define the symbols as:
| Symbol | Definition | Initial Value/Literature Value | Fitted |
|---|---|---|---|
| \([Arab:AraC]\) | The concentration of associated Arabinose and AraC | \(0\) | - |
| \([mRNA]\) | The concentration of mRNA | \(0\) | - |
| \([P]\) | The concentration of our product | \(0\) | - |
| \(\alpha\) | Translation rate | \(15ntd\: s^{-1}\)/length of sequence [6] | ? |
| \(\gamma_{1}\) | Degradation rate of mRNA | \(5.13\times10^{-4}s^{-1}\) [5] | ? |
| \(\gamma_{2}\) | Degradation rate of product | \(5.13\times10^{-4}s^{-1}\) [5] | ? |
| \(K_{max}\) | Maximal transcription rate | \(50ntd\: s^{-1}\)/length of sequence [6] | ? |
| \(K_{half}\) | Half-maximal transcription rate | \(160\mu M\) [8] | ? |
| \(n\) | Hill coefficient | \(2.65\) [3] | ? |
This table contains literature values for the parameters, found from a number of sources. Later we will fit the parameters to the experimental data found by the wet lab team. For now though we can plot the expression graphs using the literature values. This will provide an estimate to the time scales involved.
There are mutliple products being expressed using this inducer-promoter pair, each of different sequence lenghts. Here is a table showing the relevant proteins and sequence lengths:
| Product | Sequence Length (/bp) |
|---|---|
| pBAD HisB DNase DsbA | 621 |
| pBAD HisB DspB YebF | |
| pBAD HisB DspB | |
| pBAD HisB MccS | 414 |
| pBAD HisB DspB Fla | |
| pBAD HisB Art-175 DsbA | 987 |
| pBAD HisB Art-175 YebF | 1284 |
| pBAD HisB Art-E | 632 |
| pBAD HisB Art-175 Fla | 1095 |
| pBAD HisB Art-175 | 936 |
| pBAD HisB DNase | 570 |
We now can run our model of the system by solving the set of equations using the MATLAB equation solver ode15s. Below is a plot of the concentration of product against time for each protein expressed with this inducer-promoter pair where the expression is induced by a step function:
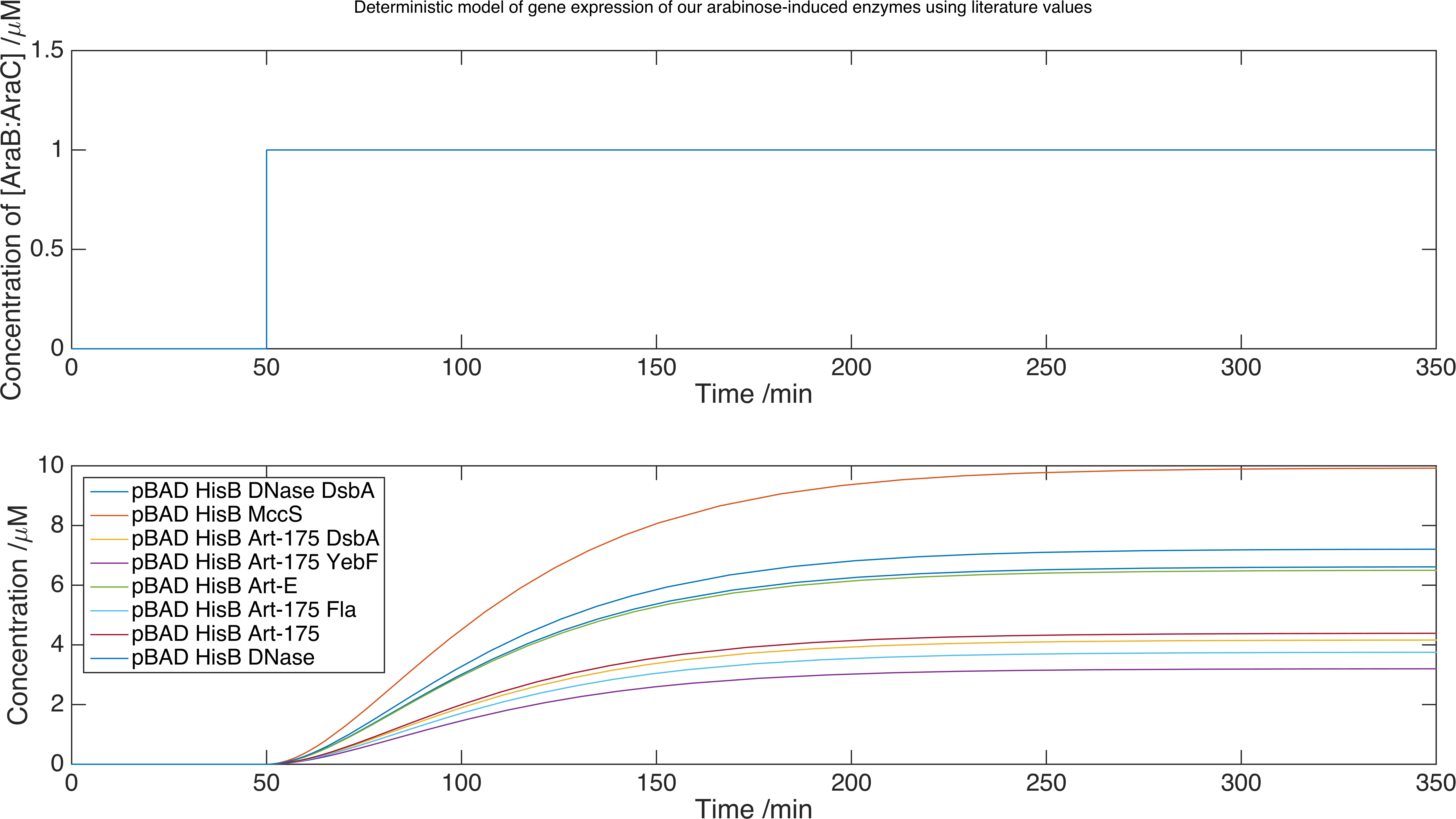
Delivery
With the information about the rates of production and concentrations of our products we can look at how the products behave once they leave the cell. This involves modelling the diffusion of the products in different topologies, each associated with a potential physical design of the catheter. With this information we can provide a better estimate of the time scale that our project is working on and assess any need for optimisation.
Dispersin B
Dispersin B is one of the anti-biofilm agents we are using in our project and will be the focus of this delivery section. As such we will assume that conclusions reached apply to all of our enzymes.
A concentration of Dispersin B of 60μg/ml is required to destroy a biofilm that has already formed on a surface. This equates to a concentration of 1.50μM. This is higher than the steady-state gene expression concentration we can expect from our cells, meaning that our system cannot rely solely on diffusion to transport our enzymes to the biofilm. We will therefore model these diffusion systems assuming that our cells are expressing at a 2μM concentration and later we will look at optimising the gene expression to this level.
Beads
Diffusion
The bead delivery system consists of our cells being contained in alginate spheres. Water is passed through the container filled with the beads allowing our enzymes to diffuse from the alginate to the required concentration. More details about the design of the system can be found here.
To determine the convection mass transfer coefficient of Dispersin B from our gel spheres we looked at the diffusion data obtained from this experiment involving the diffusion of crystal violet from our beads. By analysing the system we can produce a theoretical form for the concentration of crystal violet in the bulk water as a function of time:
\[c_{f}=\dfrac{c_{bo}}{1+\frac{V_{f}}{V_{b}}}\left(1-\exp\left(\dfrac{-K_{m}A_{b}\left(1+\frac{V_{f}}{V_{b}}\right)t}{V_{f}}\right)\right)\]| Symbol | Definition | Value | Units |
|---|---|---|---|
| \(A_{b}\) | Total surface area of the beads | \(0.0238\) | \(m^{2}\) |
| \(V_{b}\) | Total volume of beads | \(1.3463\times10^{-5}\) | \(m^{3}\) |
| \(c_{bo}\) | Initial concentration in beads | \(0.02451107\) | \(M\) |
| \(V_{f}\) | Volume of fluid surrounding the beads | \(V_{f}=V_{fo}-\dfrac{1\times10^{-6}}{10}t\) | \(m^{3}\) |
| \(V_{fo}\) | Initial volume of fluid surrounding the beads | \(1\times10^{-4}\) | \(m^{3}\) |
| \(t\) | Time | \(-\) | \(min\) |
| \(c_{f}\) | Concentration of fluid surrounding beads | \(-\) | \(M\) |
| \(K_{m}\) | Convection mass diffusion coefficient | To be fitted | \(mmin^{-1}\) |
The volume of fluid is also a function of time in order to account for the removal of 1ml of water every 10 minutes. The area and volume of the beads is that of 660 spheres with diameter 3.39mm.
However, the number of beads is an estimate. Because of this, in order to fit the curve to the experimental data we must scale the experimental data by an unknown factor. Therefore we preface our equation with an arbitrary scaling factor which, along with the convection diffusion coefficient - \(Km\), is determined by our fitting function.
Our fitting script, detailed here, returned the value of \(K_{m} = 1.7265\times 10^{-5} mmin^{-1}\).
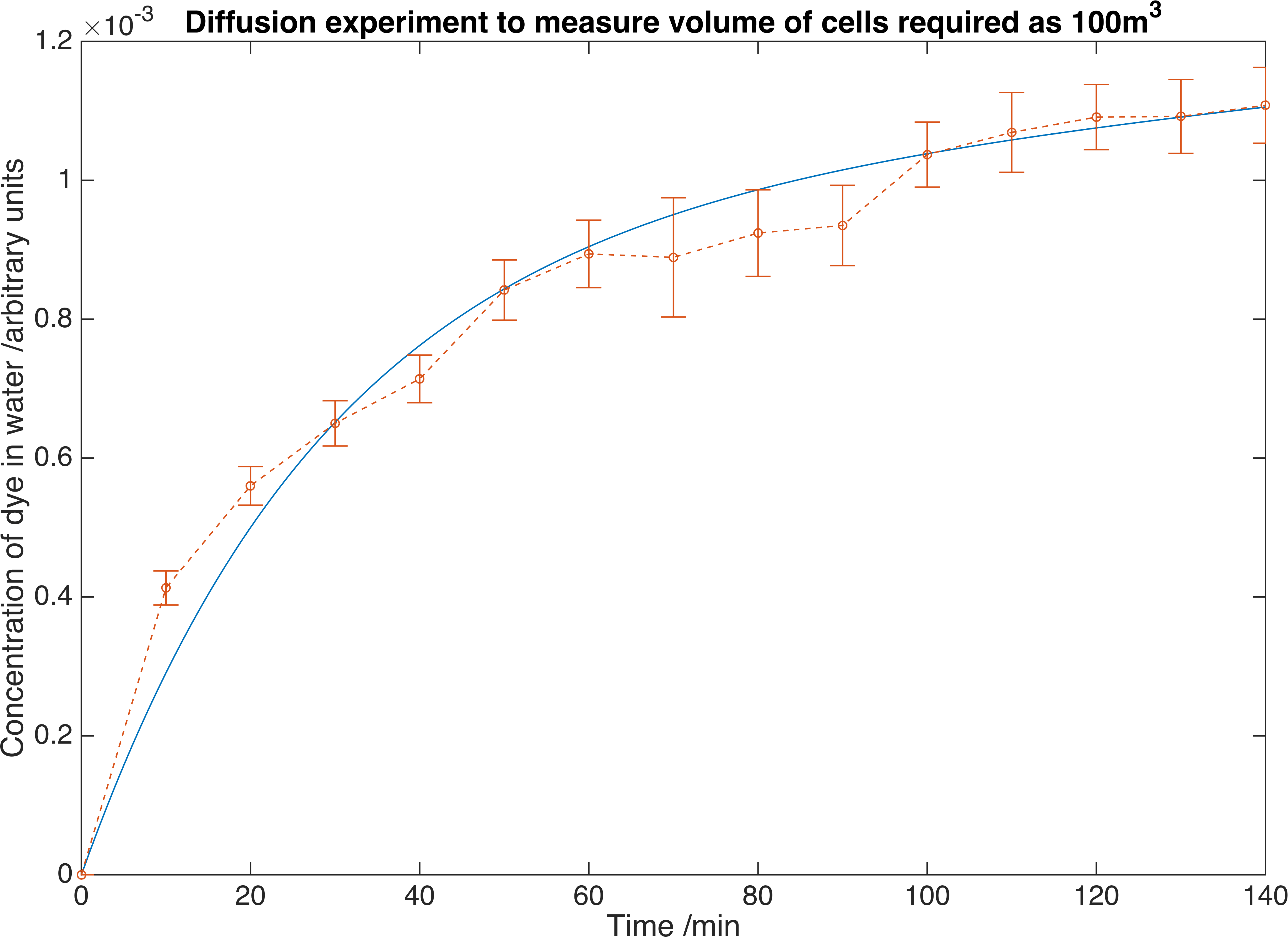
Our fitted curve plotted with the experimental data
Dispersin B is a significantly larger molecule than crystal violet so this diffusion coefficient will not be close to that for Dispersin B. To correct this we need to make use of similarity. More specifically we take the Sherwood Numbers of the systems to be equal therefore:
\[\left(\dfrac{K_{m}R}{D}\right)_{crystal violet} = \left(\dfrac{K_{m}R}{D}\right)_{Dispersin B}\]| Symbol | Definition | Value | Units |
|---|---|---|---|
| \(D_{crystal violet}\) | Mass diffusivity of crystal violet in water | \(2.8652\times10^{9}\) | \(\mu m^{2}s^{-1}\) |
| \(D_{Dispersin B}\) | Mass diffusivity of Dispersin B in water | \(100\) | \(\mu m^{2}s^{-1}\) |
| \(R\) | Radius of bead | \(1.695\) | \(mm\) |
By rearranging this we arrive at \(\left(K_{m}\right)_{DispersinB} = 6.03\times10^{-13} mmin^{-1}\)
Mass Exchange
This result allows us to theorise a mass exchange system. As a first estimate we will assume that the flow through the beads is sufficiently slow to use the convection diffusion coefficient found above. It is also assumed that the gene expression happens on a faster time scale than the diffusion from the beads to the water, enabling us to assume the concentration of enzyme in the beads remains constant. This is supported by our gene expression models. We can now visualize how the concentrations of the fluid will vary with distance along the mass exchanger:
The overall system can now be described with the equation:
\[J = K_mA\dfrac{c_{fo}-c_{fi}}{\ln\left(\dfrac{c_{B}-c_{fi}}{c_{B}-c_{fo}}\right)}\]Therefore
\[A = J\dfrac{\ln\left(\dfrac{c_{B}-c_{fi}}{c_{B}-c_{fo}}\right)}{K_{m}\left(c_{fo}-c_{fi}\right)}\]Where \(J=Q\left(c_{fo}-c_{fi}\right)\) and \(Q\) is the volume flow rate of water. We have chosen a flow rate range of 10-100ml/min as this is accepted as a safe artificial bladder fill rate. This range results in the following number of beads required to reach the desired concentration:
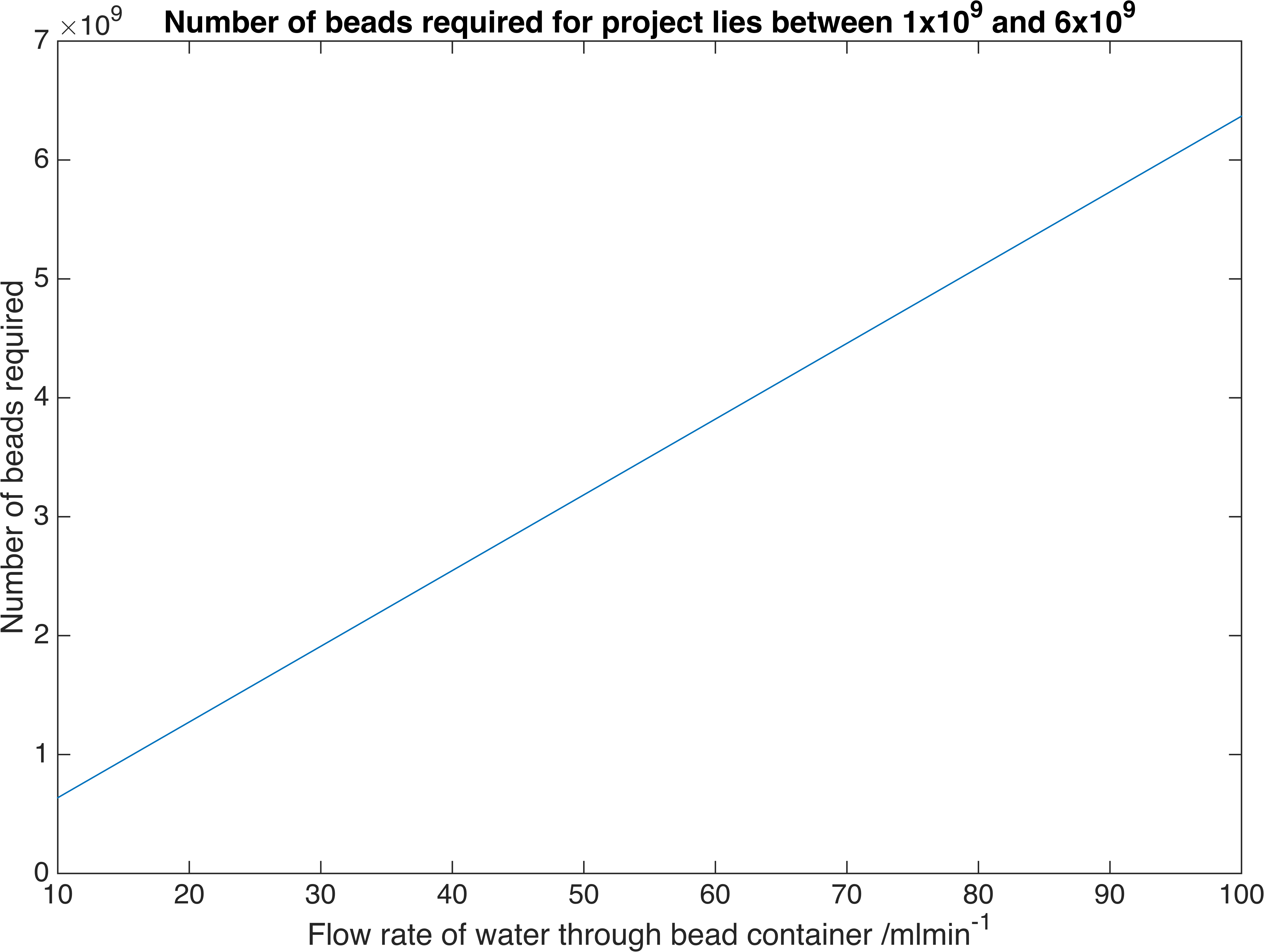
The number of beads required against the flow rate of water through the mass exchange section
Therefore a volume of between \(20.3-203m^3\) of beads is required, assuming a packing efficiency of 64%.
However, as stated earlier this estimation relies on the fluid flowing around the beads is slow enough to be approximated as stationary, meaning that mass transfer occurs as natural convection. Although there may be a very large volume of beads and a slow fluid flow rate, the area through which the fluid can flow is likely small enough that the velocity of the fluid is non-negligable.
References