Difference between revisions of "Team:Oxford/Modeling"
| Line 3: | Line 3: | ||
<html> | <html> | ||
<div class="container-fluid page-heading"> | <div class="container-fluid page-heading"> | ||
| − | <h3> | + | <h3>Modelling</h3> |
</div> | </div> | ||
<div class="container-fluid"> | <div class="container-fluid"> | ||
<div class="row"> | <div class="row"> | ||
<div class="col-md-9"> | <div class="col-md-9"> | ||
| − | <div class=" | + | <div class="slim"> |
| − | <div class=" | + | <div class="section" id="introduction"> |
<h2>Introduction</h2> | <h2>Introduction</h2> | ||
<p> | <p> | ||
| − | + | Mathematical modelling plays a crucial role in Synthetic Biology by acting as a link between the conception and the physical realisation of a biological circuit. Our modelling team has focussed on building a better picture of the project to evaluate the effectiveness of initial designs, as well as to provide insight into how the system can (or must) be improved. To do this we have split our modelling efforts into three main sections: <em>Characterising our Cells</em>, <em>Interaction with the Environment</em>, and <em>Interaction with the Biofilm</em>. By combining the information gathered in each of these section we hope to ultimately answer the question: Is our system feasible? If not, where should the design be altered? | |
</p> | </p> | ||
<p> | <p> | ||
| − | + | To help readers of all kinds and specialisations understand this page we have produced guides for all the modelling techniques used in this section which are available on the Modelling Tutorial page and will be linked to when relevant on this page. | |
</p> | </p> | ||
</div> | </div> | ||
| − | + | <div class="section-spacer"></div> | |
| − | + | <div class="section" id="characterising-our-cells"> | |
| − | + | <h2>Characterising Our Cells</h2> | |
| − | + | <p> | |
| − | < | + | In this section we look at our cells in isolation in order to assess their functionality and answer important questions such as “How long does it take to produce a certain concentration of product?”, and “What are the main limiting rates/concentrations?”. The first will help us assess the feasibility of our project, ie are our cells too slow? The second will aid us in further optimising our design. |
| − | <div id=" | + | </p> |
| − | <h3> | + | <div id="characterising-our-cells-arab"> |
| + | <h3>Arabinose Induced</h3> | ||
<p> | <p> | ||
| − | + | We have decided to use an arabinose induced promoter for the expression of a number of our proteins. This promoter can be modelled as the following chemical system: | |
</p> | </p> | ||
| − | + | \[\overset{\alpha_{1}}{\rightarrow}Arab\quad\overset{\alpha_{5}}{\rightarrow}AraC\] | |
| − | + | \[Arab+AraC\mathrel{\mathop{\rightleftharpoons}^{\mathrm{\alpha_{2}}}_{\mathrm{\alpha_{3}}}}Arab:AraC\] | |
| − | + | \[\overset{K}{\rightarrow}mRNA\overset{\alpha_{4}}{\rightarrow}P\] | |
| − | + | \[Arab\overset{\gamma_{1}}{\rightarrow}\phi\quad AraC\overset{\gamma_{2}}{\rightarrow}\phi\quad mRNA\overset{\gamma_{3}}{\rightarrow}\phi\quad P\overset{\gamma_{4}}{\rightarrow}\phi\] | |
<p> | <p> | ||
| − | + | You can find out more information about how this promoter works here. | |
| + | </p> | ||
| + | <p> | ||
| + | From this system of chemical reactions we can derive the following set of ODEs[<a href="#Ref4">4</a>]: | ||
</p> | </p> | ||
\[\dfrac{d\left[Arab\right]}{dt}=\alpha_{1}+\alpha_{2}\left[Arab\right]\left[AraC\right]-\alpha_{3}\left[Arab:AraC\right]-\gamma_{1}\left[Arab\right]\] | \[\dfrac{d\left[Arab\right]}{dt}=\alpha_{1}+\alpha_{2}\left[Arab\right]\left[AraC\right]-\alpha_{3}\left[Arab:AraC\right]-\gamma_{1}\left[Arab\right]\] | ||
| Line 42: | Line 46: | ||
\[\dfrac{d\left[Arab:AraC\right]}{dt}=\alpha_{3}\left[Arab:AraC\right]-\alpha_{2}\left[Arab\right]\left[AraC\right]\] | \[\dfrac{d\left[Arab:AraC\right]}{dt}=\alpha_{3}\left[Arab:AraC\right]-\alpha_{2}\left[Arab\right]\left[AraC\right]\] | ||
| − | \[\dfrac{d\left[AraC\right]}{dt}=\alpha_{5} | + | \[\dfrac{d\left[AraC\right]}{dt}=\alpha_{5}-\alpha_{2}\left[Arab\right]\left[AraC\right]+\alpha_{3}\left[Arab:AraC\right]-\gamma_{2}\left[AraC\right]\] |
\[\dfrac{d[mRNA]}{dt}=K_{max}\dfrac{[Arab:AraC]^{n}}{K_{half}^{n}+[Arab:AraC]^{n}}-\gamma_{3}[mRNA]\] | \[\dfrac{d[mRNA]}{dt}=K_{max}\dfrac{[Arab:AraC]^{n}}{K_{half}^{n}+[Arab:AraC]^{n}}-\gamma_{3}[mRNA]\] | ||
| Line 48: | Line 52: | ||
\[\dfrac{d\left[P\right]}{dt}=\alpha_{4}\left[mRNA\right]-\gamma_{4}\left[P\right]\] | \[\dfrac{d\left[P\right]}{dt}=\alpha_{4}\left[mRNA\right]-\gamma_{4}\left[P\right]\] | ||
<p> | <p> | ||
| − | Where we define the symbols | + | Where we define the symbols as: |
| − | <table | + | <table class="table table-striped"> |
| − | < | + | <thead> |
<th>Symbol</th> | <th>Symbol</th> | ||
<th>Definition</th> | <th>Definition</th> | ||
<th>Initial Value/Literature Value</th> | <th>Initial Value/Literature Value</th> | ||
<th>Fitted</th> | <th>Fitted</th> | ||
| − | </ | + | </thead> |
<tr> | <tr> | ||
<td>\([Arab]\)</td> | <td>\([Arab]\)</td> | ||
<td>The concentration of Arabinose</td> | <td>The concentration of Arabinose</td> | ||
| − | <td>\(1\times10^{- | + | <td>\(1\times10^{-5}M\)</td> |
<td>-</td> | <td>-</td> | ||
</tr> | </tr> | ||
| Line 161: | Line 165: | ||
</p> | </p> | ||
<p> | <p> | ||
| − | + | This table contains literature values for the parameters, found from a number of sources. Later we will fit the parameters to the experimental data found by the wet lab team. For now though we can plot the expression graphs using the literature values. This will provide an estimate to the time scales involved. | |
</p> | </p> | ||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
<p> | <p> | ||
| − | + | There are mutliple products being expressed using this inducer-promoter pair, each of different sequence lenghts. Here is a table showing the relevant proteins and sequence lengths: | |
</p> | </p> | ||
| − | < | + | <table class="table table-striped"> |
| − | < | + | <thead> |
| − | < | + | <tr> |
| − | < | + | <th> |
| + | Product | ||
| + | </th> | ||
| + | <th> | ||
| + | Sequence Length (/bp) | ||
| + | </th> | ||
| + | </tr> | ||
| + | </thead> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB DNase DsbA | ||
| + | </td> | ||
| + | <td> | ||
| + | 621 | ||
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB DspB YebF | ||
| + | </td> | ||
| + | <td> | ||
| − | < | + | </td> |
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB DspB | ||
| + | </td> | ||
| + | <td> | ||
| − | < | + | </td> |
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB MccS | ||
| + | </td> | ||
| + | <td> | ||
| + | 414 | ||
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB DspB Fla | ||
| + | </td> | ||
| + | <td> | ||
| − | < | + | </td> |
| − | + | </tr> | |
| − | < | + | <tr> |
| − | + | <td> | |
| − | < | + | pBAD HisB Art-175 DsbA |
| − | </ | + | </td> |
| − | + | <td> | |
| − | </ | + | 987 |
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB Art-175 YebF | ||
| + | </td> | ||
| + | <td> | ||
| + | 1284 | ||
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB Art-E | ||
| + | </td> | ||
| + | <td> | ||
| + | 632 | ||
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB Art-175 Fla | ||
| + | </td> | ||
| + | <td> | ||
| + | 1095 | ||
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB Art-175 | ||
| + | </td> | ||
| + | <td> | ||
| + | 936 | ||
| + | </td> | ||
| + | </tr> | ||
| + | <tr> | ||
| + | <td> | ||
| + | pBAD HisB DNase | ||
| + | </td> | ||
| + | <td> | ||
| + | 570 | ||
| + | </td> | ||
| + | </tr> | ||
| + | </table> | ||
<p> | <p> | ||
| − | + | We now can run our model of the system by solving the set of equations using the MATLAB equation solver ode15s. Below is a plot of the concentration of product against time for each protein expressed with this inducer-promoter pair: | |
| − | + | ||
</p> | </p> | ||
| + | <div class="image image-full"> | ||
| + | <img src="https://static.igem.org/mediawiki/2015/f/f6/Ox_arab_induced_proteins.png"/> | ||
| + | </div> | ||
</div> | </div> | ||
</div> | </div> | ||
| − | + | <div class="section-spacer"></div> | |
| − | + | <div class="section" id="interaction-with-environment"> | |
| − | + | <h2>Interaction with the Environment</h2> | |
| − | + | <p> | |
| − | < | + | With the information about the rates of production and concentrations of our products we can look at how the products behave once they leave the cell. This involves modelling the diffusion of the products in different topologies, each associated with a potential physical design of the catheter. With this information we can provide a better estimate of the time scale that our project is working on and assess any need for optimisation. |
| + | </p> | ||
| + | <p> | ||
| + | In addition, one of our systems relies on the detection of the biofilm to cause lysis. We will look at the how the quorum sensing signal moves to the cells. | ||
| + | </p> | ||
</div> | </div> | ||
| − | <div | + | <div class="section-spacer"></div> |
| − | + | <div class="section" id="interaction-with-biofilm"> | |
| − | + | <h2>Interaction with the Biofilm</h2> | |
| − | + | <p> | |
| − | + | Once the antibiofilm agents have arrived at the biofilm, what impact do they have? We will look into the required concentrations and rates of the antibiofilm agents at the biofilm to overcome its rate of growth. | |
| − | + | </p> | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
</div> | </div> | ||
| − | <div | + | <div class="section-spacer"></div> |
| − | + | <div class="section" id="overall"> | |
| − | + | <h2>Combined: What do we know about the system as a whole?</h2> | |
| − | + | <p> | |
| − | + | What we write here depends on the results of the previous section. | |
| − | + | </p> | |
| − | + | <div id="overall-time"> | |
| − | + | <h3>How quickly will the system run?</h3> | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | < | + | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
</div> | </div> | ||
| − | + | <div id="overall-quantity"> | |
| − | + | <h3>How many engineered bacteria do we need?</h3> | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | <h3> | + | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
</div> | </div> | ||
| − | + | <div id="overall-concentration"> | |
| − | + | <h3>What is the minimum steady state concentration we need to reach to overcome the growth rate of the biofilm?</h3> | |
| − | + | ||
| − | <h3> | + | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
</div> | </div> | ||
</div> | </div> | ||
| Line 370: | Line 314: | ||
<div class="section" id="references"> | <div class="section" id="references"> | ||
<h2>References</h2> | <h2>References</h2> | ||
| − | < | + | <p> |
| − | + | </p> | |
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
| − | + | ||
</div> | </div> | ||
</div> | </div> | ||
| Line 395: | Line 324: | ||
</li> | </li> | ||
<li> | <li> | ||
| − | <a href="# | + | <a href="#characterising-our-cells">Characterising Our Cells</a> |
<ul class="nav nav-stacked"> | <ul class="nav nav-stacked"> | ||
| − | <li><a href="# | + | <li><a href="#characterising-our-cells-arab">Arabinose Induced</a></li> |
| − | + | ||
</ul> | </ul> | ||
</li> | </li> | ||
<li> | <li> | ||
| − | <a href="# | + | <a href="#interaction-with-environment">Interaction with the Environment</a> |
| − | + | </li> | |
| − | + | <li> | |
| − | + | <a href="#interaction-with-biofilm">Interaction with the Biofilm</a> | |
| − | + | ||
</li> | </li> | ||
<li> | <li> | ||
| − | <a href="# | + | <a href="#overall">Combined: What do we know about the system as a whole?</a> |
<ul class="nav nav-stacked"> | <ul class="nav nav-stacked"> | ||
| − | <li><a href="# | + | <li><a href="#overall-time">How quickly will the system run?</a></li> |
| − | <li><a href="# | + | <li><a href="#overall-quantity">How many engineered bacteria do we need?</a></li> |
| − | <li><a href="# | + | <li><a href="#overall-concentration">What is the minimum steady state concentration we need to reach to overcome the growth rate of the biofilm?</a></li> |
</ul> | </ul> | ||
</li> | </li> | ||
Revision as of 12:57, 4 September 2015
Modelling
Introduction
Mathematical modelling plays a crucial role in Synthetic Biology by acting as a link between the conception and the physical realisation of a biological circuit. Our modelling team has focussed on building a better picture of the project to evaluate the effectiveness of initial designs, as well as to provide insight into how the system can (or must) be improved. To do this we have split our modelling efforts into three main sections: Characterising our Cells, Interaction with the Environment, and Interaction with the Biofilm. By combining the information gathered in each of these section we hope to ultimately answer the question: Is our system feasible? If not, where should the design be altered?
To help readers of all kinds and specialisations understand this page we have produced guides for all the modelling techniques used in this section which are available on the Modelling Tutorial page and will be linked to when relevant on this page.
Characterising Our Cells
In this section we look at our cells in isolation in order to assess their functionality and answer important questions such as “How long does it take to produce a certain concentration of product?”, and “What are the main limiting rates/concentrations?”. The first will help us assess the feasibility of our project, ie are our cells too slow? The second will aid us in further optimising our design.
Arabinose Induced
We have decided to use an arabinose induced promoter for the expression of a number of our proteins. This promoter can be modelled as the following chemical system:
\[\overset{\alpha_{1}}{\rightarrow}Arab\quad\overset{\alpha_{5}}{\rightarrow}AraC\] \[Arab+AraC\mathrel{\mathop{\rightleftharpoons}^{\mathrm{\alpha_{2}}}_{\mathrm{\alpha_{3}}}}Arab:AraC\] \[\overset{K}{\rightarrow}mRNA\overset{\alpha_{4}}{\rightarrow}P\] \[Arab\overset{\gamma_{1}}{\rightarrow}\phi\quad AraC\overset{\gamma_{2}}{\rightarrow}\phi\quad mRNA\overset{\gamma_{3}}{\rightarrow}\phi\quad P\overset{\gamma_{4}}{\rightarrow}\phi\]You can find out more information about how this promoter works here.
From this system of chemical reactions we can derive the following set of ODEs[4]:
\[\dfrac{d\left[Arab\right]}{dt}=\alpha_{1}+\alpha_{2}\left[Arab\right]\left[AraC\right]-\alpha_{3}\left[Arab:AraC\right]-\gamma_{1}\left[Arab\right]\] \[\dfrac{d\left[Arab:AraC\right]}{dt}=\alpha_{3}\left[Arab:AraC\right]-\alpha_{2}\left[Arab\right]\left[AraC\right]\] \[\dfrac{d\left[AraC\right]}{dt}=\alpha_{5}-\alpha_{2}\left[Arab\right]\left[AraC\right]+\alpha_{3}\left[Arab:AraC\right]-\gamma_{2}\left[AraC\right]\] \[\dfrac{d[mRNA]}{dt}=K_{max}\dfrac{[Arab:AraC]^{n}}{K_{half}^{n}+[Arab:AraC]^{n}}-\gamma_{3}[mRNA]\] \[\dfrac{d\left[P\right]}{dt}=\alpha_{4}\left[mRNA\right]-\gamma_{4}\left[P\right]\]Where we define the symbols as:
| Symbol | Definition | Initial Value/Literature Value | Fitted |
|---|---|---|---|
| \([Arab]\) | The concentration of Arabinose | \(1\times10^{-5}M\) | - |
| \([AraC]\) | The concentration of AraC | \(1\times10^{-5}M\) | - |
| \([Arab:AraC]\) | The concentration of associated Arabinose and AraC | \(0\) | - |
| \([mRNA]\) | The concentration of mRNA | \(0\) | - |
| \([P]\) | The concentration of our product | \(0\) | - |
| \(\alpha_{1}\) | Basal production of Arabinose | ??? | ? |
| \(\alpha_{2}\) | Association constant | \(2.8\times10^{7}s^{-1}\) [7] | ? |
| \(\alpha_{3}\) | Dissociation constant | \(0.022s^{-1}\) [7] | ? |
| \(\alpha_{4}\) | Translation rate | \(15ntd\: s^{-1}\)/length of sequence [6] | ? |
| \(\alpha_{5}\) | Basal production of AraC | ??? | ? |
| \(\gamma_{1}\) | Degradation rate of Arabinose | \(5.13\times10^{-4}s^{-1}\) [5] | ? |
| \(\gamma_{2}\) | Degradation rate of AraC | \(5.13\times10^{-4}s^{-1}\) [5] | ? |
| \(\gamma_{3}\) | Degradation rate of mRNA | \(5.13\times10^{-4}s^{-1}\) [5] | ? |
| \(\gamma_{4}\) | Degradation rate of product | \(5.13\times10^{-4}s^{-1}\) [5] | ? |
| \(K_{max}\) | Maximal transcription rate | \(50ntd\: s^{-1}\)/length of sequence [6] | ? |
| \(K_{half}\) | Half-maximal transcription rate | \(160\mu M\) [8] | ? |
| \(n\) | Hill coefficient | \(2.65\) [3] | ? |
This table contains literature values for the parameters, found from a number of sources. Later we will fit the parameters to the experimental data found by the wet lab team. For now though we can plot the expression graphs using the literature values. This will provide an estimate to the time scales involved.
There are mutliple products being expressed using this inducer-promoter pair, each of different sequence lenghts. Here is a table showing the relevant proteins and sequence lengths:
| Product | Sequence Length (/bp) |
|---|---|
| pBAD HisB DNase DsbA | 621 |
| pBAD HisB DspB YebF | |
| pBAD HisB DspB | |
| pBAD HisB MccS | 414 |
| pBAD HisB DspB Fla | |
| pBAD HisB Art-175 DsbA | 987 |
| pBAD HisB Art-175 YebF | 1284 |
| pBAD HisB Art-E | 632 |
| pBAD HisB Art-175 Fla | 1095 |
| pBAD HisB Art-175 | 936 |
| pBAD HisB DNase | 570 |
We now can run our model of the system by solving the set of equations using the MATLAB equation solver ode15s. Below is a plot of the concentration of product against time for each protein expressed with this inducer-promoter pair:
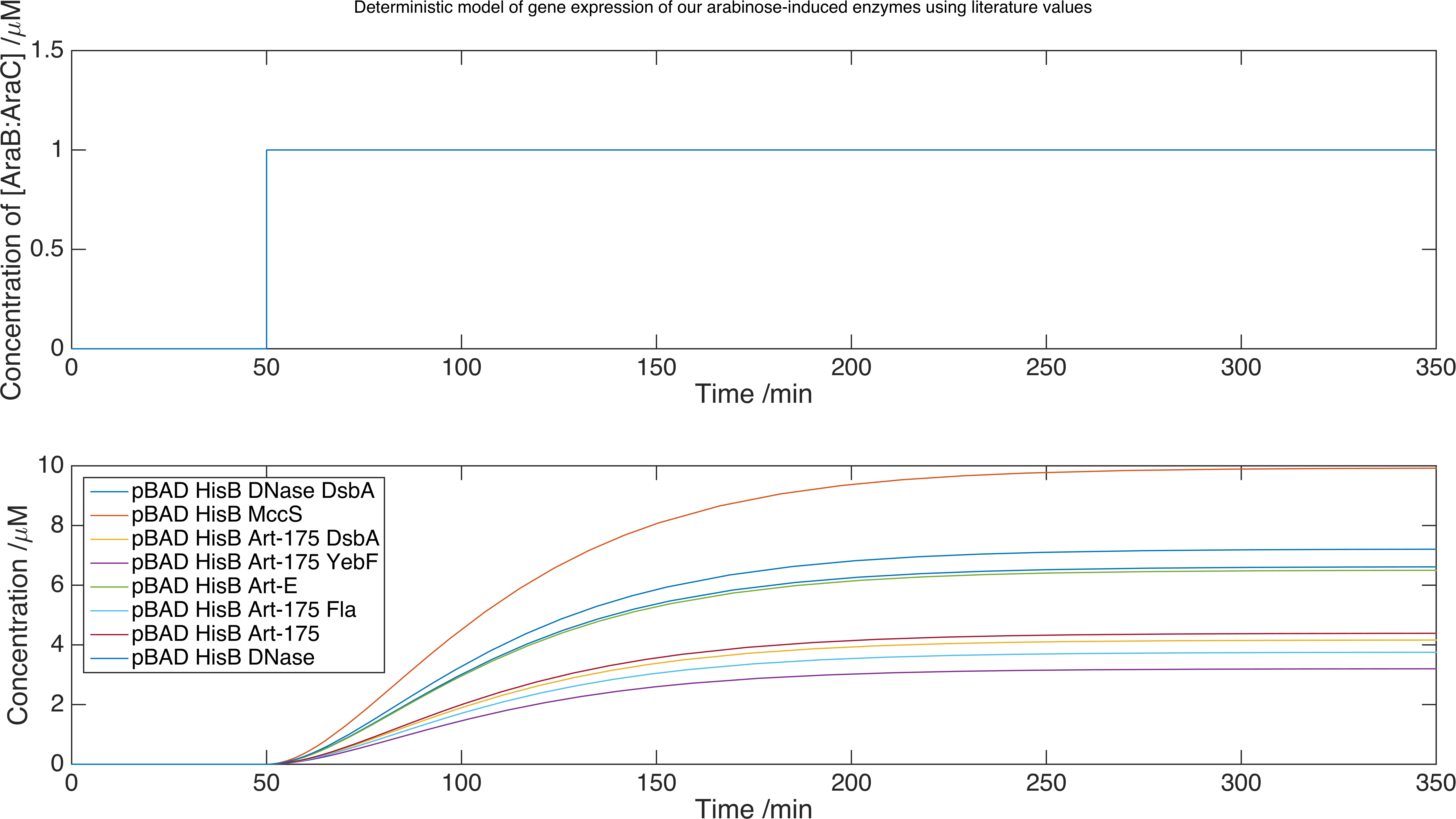
Interaction with the Environment
With the information about the rates of production and concentrations of our products we can look at how the products behave once they leave the cell. This involves modelling the diffusion of the products in different topologies, each associated with a potential physical design of the catheter. With this information we can provide a better estimate of the time scale that our project is working on and assess any need for optimisation.
In addition, one of our systems relies on the detection of the biofilm to cause lysis. We will look at the how the quorum sensing signal moves to the cells.
Interaction with the Biofilm
Once the antibiofilm agents have arrived at the biofilm, what impact do they have? We will look into the required concentrations and rates of the antibiofilm agents at the biofilm to overcome its rate of growth.
Combined: What do we know about the system as a whole?
What we write here depends on the results of the previous section.
How quickly will the system run?
How many engineered bacteria do we need?
What is the minimum steady state concentration we need to reach to overcome the growth rate of the biofilm?
References