Team:Birkbeck/Composite Part




Our BioBricks
Composite Parts
tfa (tail fibre assembly) gene circuit (BBa_K1846001)

This part provides the gene sequence for tfa (tail fibre assembly) protein, together with a TetR repressible promoter (TetO, available separately as BioBrick part BBa_R0040), ribosome binding site (available separately as part BBa_B0034) and an rrNB T1 terminator (available separately as BioBrick part BBa_B0010).
The tfa (tail fibre assembly) protein of bacteriophage lambda assists in the assembly of the stf (short tail fibre) protein into a functional short tail fibre [1],[2]. Tfa gene displays a high level of homology (~40%) with the gp38 of bacteriophage T4 [2],[3],[4].
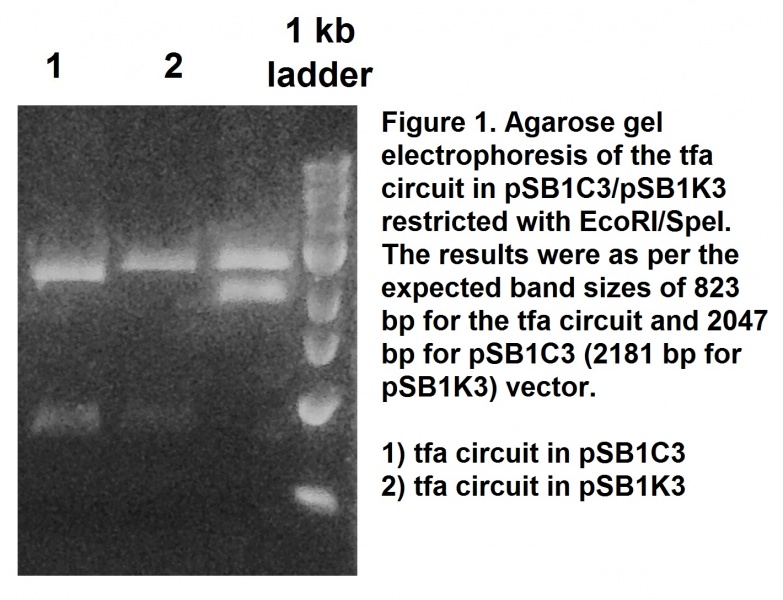
This tfa circuit was synthesised removing the illegal restriction sites to make the sequence BioBrick-compatible. The sequence was cloned into a pSB1C3 and the success of the cloning procedure was confirmed by restriction with EcoRI and SpeI restriction enzymes and followed by agarose gel electrophoresis (Figure 1). The construct was also confirmed by sequencing. This BioBrick was registered as part BBa_K1846001.
TetR circuit (BBa_K1846003)

This is a constitutive part for the production of the tetracyline repressor TetR, using a constitutive, medium-copy chloramphenicol promoter (BBa_I14033), ribosome binding sequence (BBa_B0031) and double terminator (rrnb T1 terminator followed by a T7Te terminator BBa_B0010).
TetR generator cassette BBa_P0140 and P(Cat) promoter BBa_I14033 were obtained from 2015 iGEM distribution kit and cloned into a single pSB1C3 vector. Correct cloning of the parts into the pSB1C3 shipping vector was confirmed by agarose gel electrophoresis (Figure 2) and Sanger sequencing. This BioBrick has been registered as part BBa_K1846003.
TetR-regulated tfa (tail fibre assembly) circuit (BBa_K1846007)

To control production of the tail fibre assembly protein to prevent toxicity to the cell, we have combined the TetR circuit (BBa_K1846003) and the tfa circuit (BBa_K1846001) into a single BioBrick. With the production of the tfa protein under control of a Tet-repressible promoter, the coding sequence will remain inactive due to production of TetR. However, on addition of anhydrous tetracycline, TetR will preferably bind to this molecule, allowing the initiation of transcription. The correct cloning of this BioBrick was confirmed through Sanger sequencing and agarose gel electrophoresis (see results). This BioBrick has been registered as part BBa_K1846007.
We characterised this construct by analysing the soluble protein fraction of the cell lysate (Figure 1,2). Under the TetR regulation, tfa gene is expressed under a tight control, and can thus be visualised on the gel, 194 aa, MW: 21602.2 Da, Figure 1, samples 7,8,9 & 11,12 and 13; Figure 2, samples 4,5 and 6. Tfa gene circuit does not produce tfa protein bands, Figure 1, samples 1, 2 and 3. We attribute the lack of bands in these samples to protein overexpression and aggregation into insoluble fraction. Under regulation of TetR, even when induced, the expression of tfa is considerably lower than when unregulated and hence, we hypothesise, a considerable amount of soluble material can be recovered. Our next step is to analyse the cell pellet to determine the presence of the tfa gene in the insoluble fraction.


cI-Cro circuit (BBa_K1846005)

We have created a regulatory circuit to control the lysogenic and lytic phases of bacteriophage lambda. The cI-Cro construct contains a circuit for the production - via a constitutive promoter - of the cI repressor protein (also known as the Lambda Repressor), which is responsible for keeping bacteriophage lambda in the lysogenic cycle through the cooperative binding of two repressor dimers to the DNA, repressing the Cro gene. The circuit uses a T7 promoter to drive the expression of the Cro gene in the opposite direction to the cI gene, essentially silensing the expression of the cI gene. The strength of the promoter used means that in the presence of T7 DNA polymerase, production of the Cro repressor will incapacitate production of the cI repressor and thus enables the switch from lysogenic cycle to the lytic cycle. The correct cloning of this BioBrick was confirmed through Sanger sequencing and agarose gel electrophoresis (see results). This BioBrick has been registered as part BBa_K1846005.
We characterised this construct by analysing the soluble protein fraction of the cell lysate (Figure 3). As cI protein is produced constitutively in our circuit, we expected to see a cI protein band at ca 26 kDa (cI = 237 aa, MW: 26211.8 Da). Our control E.coli 10β cells containing the cI/cro construct are showing the expected bands at ca 26 kDa, Figure 3, samples 7, 8 and 9. We attribute lack of cI bands in our T7 Express cell lysate due to T7 RNAP leakage, which would silence the cI expression. This hypothesis requires further investigation, however our preliminary results suggest the circuit could be functional.

[1] Hashemolhosseini, S., Stierhof, Y. D., Hindennach, I., & Henning, U. (1996). Characterization of the helper proteins for the assembly of tail fibers of coliphages T4 and λ. Journal of Bacteriology, 178(21), 6258–6265.
[2] Montag, D., Schwarz, H., & Henning, U. (1989). A component of the side tail fiber of E. coli bacteriophage λ can functionally replace the receptor-recognizing part of a long tail fiber protein of the unrelated bacteriophage T4. Journal of Bacteriology, 171(8), 4378–4384.
[3] Hendrix, R. W., & Duda, R. L. (1992). Bacteriophage lambda PaPa: not the mother of all lambda phages. Science (New York, N.Y.), 258(5085), 1145–1148. doi:10.1126/science.1439823
[4] Haggard-Ljungquist, E., Halling, C., & Calendar, R. (1992). DNA sequences of the tail fiber genes of bacteriophage P2: Evidence for horizontal transfer of tail fiber genes among unrelated bacteriophages. Journal of Bacteriology, 174(5), 1462–1477.