Team:KU Leuven/Modeling/Hybrid

The hybrid model
The hybrid model represents an intermediate level of detail in between the colony level model and the internal model. Bacteria are treated as individual agents that behave according to the Keller-Segel type discretized stochastic differential equations, while chemical species are modeled using partial differential equations.
Introduction
Spatial reaction-diffusion models that rely on Partial Differential Equations (PDEs) are based on the assumption that the collective behavior of individual entities, such as molecules or bacteria, can be abstracted to the behavior of a continuous field that represents the density of those entities. The brownian motion of molecules, for instance, is the result of inherently stochastic processes that take place at the individual molecule level, but is modeled at the density level by Fick’s laws of diffusion. These PDE-based models provide a robust method to predict the evolution of large-scale systems, but are only valid when the spatiotemporal scale is sufficiently large to neglect small-scale stochastic fluctuations and physical granularity. Moreover, such a continuous field approximation can only be made if the behavior of the individual entities is well described.
Agent-based Models
Agent-based models on the other hand explicitly treat the entities as individual “agents” that behave according to a set of “agent rules”. An agent is an object that acts independently from other agents and is influenced only by its local environment. The goal in agent-based models is to study the emergent systems-level properties of a collection of individual agents that follow relatively simple rules. In smoothed particle hydrodynamics for example, fluids are simulated by calculating the trajectory of each individual fluid particle at every timestep. Fluid properties such as the momentum at a certain point can then be sampled by taking a weighted sum of the momenta of the surrounding fluid particles. A large advantage of agent-based models is that the agent rules are arbitrarily complex and thus they allow us to model systems that do not correspond to any existing or easily derivable PDE model. However, because every agent is stored in memory and needs to be processed individually, simulating agent-based models can be computationally intensive.
Model Description
Hybrid model
The main protagonists in our pattern-forming system are cell types A and B, AHL and leucine. Cells A produce AHL as well as leucine. They are unaffected by leucine, while cells B are repelled by leucine. AHL modulates the motility of both cell types A and B, but in opposite ways. High concentrations of AHL will render cell type A unable to swim but will activate cell type B’s motility. Conversely, low concentrations of AHL causes swimming of cell type A and incessant tumbling (thus immobility) of cell type B. Lastly, cells A express the adhesin membrane protein, which causes them to stick to each other. Simply said, our system should produce spots of immobile, sticky groups of A type cells, surrounded by rings of B type cells. Any cell that finds itself outside of the region that it should be in, is able to swim to their correct place and becomes immobile there. More details can be found in the research section.
Partial Differential Equations
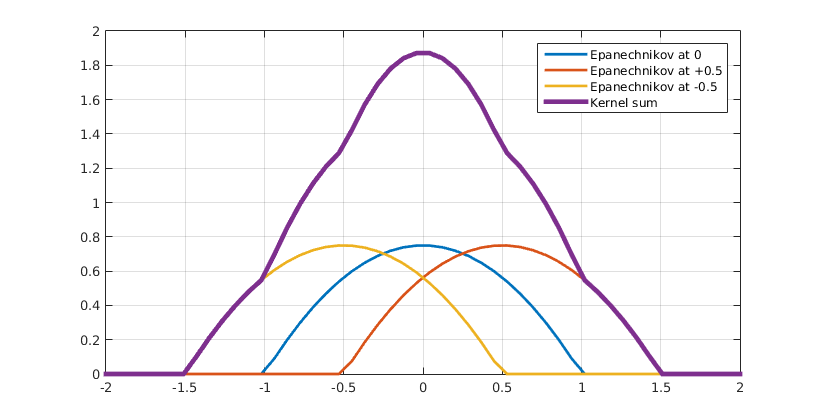
As discussed in the previous paragraph, our hybrid model incorporates chemical species using PDEs. In our system these are AHL and leucine. The diffusion of AHL and leucine can be described by the heat equation (Eq. 1). $$\frac{\partial C(\vec{r},t)}{\partial t}=\nabla^2 C(\vec{r},t) \;\;\; \text(1)$$ By using (Eq. 1) we assume that the diffusion speed is isotropic, i.e. the same in all spatial directions. This also explains why it is called the heat equation, since heat diffuses equally fast in all directions. A detailed explanation of the heat equation can be found in box 1. The second factor that needs be taken into account is the production of AHL and leucine by type A bacteria. In principle, AHL and leucine production is dependent on the dynamically-evolving internal states of all cells of type A. However, for our hybrid model we ignored the inner life of all bacteria and instead assumed that AHL and leucine production is directly proportional to the density of A type cells (Eq. 2). $$ \frac{\partial C(\vec{r},t)}{\partial t}=\alpha \cdot \rho_A(\vec{r},t) \;\;\; \text{(2)}$$ In the last paragraph we will reconsider this assumption and assign each cell an internal model. Finally, AHL and leucine are organic molecules and like most organic molecules they will degrade over time. $$ \frac{\partial C_{AHL}(\vec{r},t)}{\partial t}=-k_{AHL}\cdot C_{AHL}(\vec{r},t) \;\;\; \text{(3a)} $$ $$ \frac{\partial C_{leucine}(\vec{r},t)}{\partial t}=-k_{leucine}\cdot C_{leucine}(\vec{r},t) \;\;\; \text{(3b)} $$ We assume first-order kinetics meaning that the rate at which AHL and similarly leucine disappear is proportional to their respective concentrations (Eq. 3a and 3b) assuming neutral pH (citation). $$ \frac{\partial C(\vec{r},t)}{\partial t}=\nabla^2 C(\vec{r},t)+\alpha \cdot \rho_A(\vec{r},t)-k\cdot C(\vec{r},t) \;\;\; \text{(4)} $$ Putting it all together, we obtain (Eq. 4), both for AHL and leucine. Note that these equations have exactly the same form as the equations for AHL and leucine in the colony level model. The crucial difference however lies in the calculation of the density of cells of type A. In contrast to the colony level model, the cell density is not calculated explicitly with a PDE and is therefore not trivially known. Therefore a method to extract a density field from a spatial distribution of agents is necessary. This is addressed in the subparagraph below on coupling.
Equation of motion
Now we are ready to construct the equation of motion for cell type A and B as a superposition of the Keller-Segel SDE (Eq. 6) and the cell interaction forces, yielding (Eq. 11). $$ d\vec{r}_{A_i}(t)= \sqrt{2 \cdot \mu_A}\cdot d\vec{W} + \frac{1}{\gamma}\cdot\Bigg( \sum^{A \backslash \{ A_i\}}_j \frac{dE_{p,attraction}(r_{ij}(t))}{dr_{ij}}\cdot \vec{e}_{ij}+\sum^{B}_j \frac{dE_{p,repulsion}(r_{ij}(t))}{dr_{ij}}\cdot \vec{e}_{ij} \Bigg)\cdot dt \;\;\; (11a) $$ $$ d\vec{r}_{B_i}(t)= \chi(L(\vec{r},t),H(\vec{r},t)) \cdot \nabla L(\vec{r},t)\cdot dt + \sqrt{2 \cdot \mu_B(H(\vec{r},t))}\cdot d\vec{W} + $$ $$ \frac{1}{\gamma}\cdot\Bigg( \sum^{A\cup B\backslash \{ B_i\}}_j \frac{dE_{p,repulsion}(r_{ij}(t))}{dr_{ij}}\cdot \vec{e}_{ij} \Bigg)\cdot dt $$ $$ \chi(L(\vec{r},t),H(\vec{r},t))= \mu_{B}(H(\vec{r},t)) \cdot \frac{\kappa}{L(\vec{r},t)} $$ $$ \mu_A(H(\vec{r},t))=\left\{\begin{matrix}\mu_{A,high} & H(\vec{r},t) < H_{A,threshold}\\ \mu_{A,low} & H(\vec{r},t) \geq H_{A,threshold}\end{matrix}\right. $$ $$ \mu_B(H(\vec{r},t))=\left\{\begin{matrix} \mu_{B,high} & H(\vec{r},t) < H_{B,threshold}\\ \mu_{B,low} & H(\vec{r},t) \geq H_{B,threshold} \end{matrix}\right. \;\;\; \text{(11b)} $$ Bacteria of type A are not attracted nor repelled by leucine, so the chemotactic term falls away. All cell-cell forces are summed up to find a net force, taking into account the two different potentials due to the different interaction types. As discussed before, this net force times a constant yields the velocity due to that force, which is then multiplied by dt to obtain the displacement. For B type cells, the chemotactic term models the repulsive chemotaxis away from leucine. The chemotactic sensitivity function has a negative sign signifying that B type cells are repelled by leucine. The cell interaction term in this case is simpler because B type cells only interact repulsively. Note that the diffusion coefficient of cell types A and B switches based on the local concentration of AHL relative to a threshold AHL value, which simulates the dependency of cellular motility on AHL. The agent-based module is now fully defined but one crucial issue was skipped: AHL and leucine concentrations are calculated using PDEs and are therefore only known at grid points. Agents on the other hand reside in the space between grid points and require local concentrations as inputs to calculate their next step. This problem is part of the coupling aspect in our hybrid modelling framework and is discussed below.
Heat equation
The left-hand side of (1) is the rate of accumulation of a chemical and the right-hand side is the second spatial derivative of its concentration field. The equation can be understood by considering a one-dimensional concentration profile: if (box fig1a) the concentration is higher at both sides of a certain point, it can be approximated as a convex parabolic function. Its second derivative is positive and therefore the rate of accumulation is positive (i.e. more accumulation). If on the other hand (box fig1b) the concentration is lower on both sides of a certain point, a concave function is the correct approximation. The second derivative is therefore negative and the rate of accumulation as well (i.e. depletion). Lastly it is also possible that a lower concentration is found on one side and a higher concentration on the other. In general, the rate of accumulation (accumulation or depletion) depends on the relative differences in concentration between a point and its environment. A special case occurs when the concentration profile takes on a linear form (box fig1c). This means that the differences on both sides of the point are equal in magnitude.
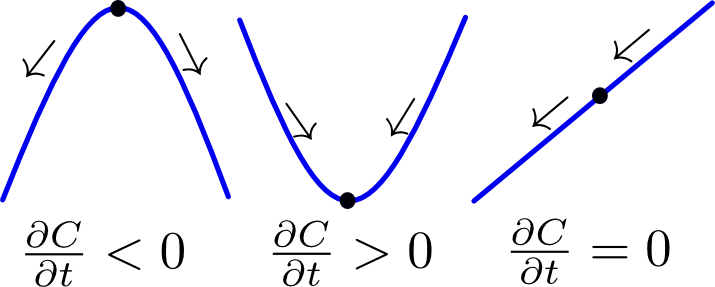
Figure 1 Illustration of the heat euqation. Click to enlarge
Life at low Reynolds number
An important parameter in fluid mechanics is the dimensionless Reynolds number, defined as the ratio of inertial forces to viscous forces, (Eq box 2.1). When this number is low, viscous forces dominate so that any acceleration phase is negligible. An important observation with regards to bacteria is that a, the characteristic dimension, is on the order of µm, which is relatively small. Filling in the other variables with the values $v ~ 20 \mu m/s $, $ \rho = 1 000 kg/m^3$ and $\eta = 0.001 Pa \cdot s $ yields a Reynolds number of around $10^{-4}$, which is very low. Due to their small size, water is viscous like honey to micro-organisms. To demonstrate this, we calculate how long it takes for a cell to slow down in water. Assuming Stokes friction, the equation of motion is (Eq box 2.2).