Difference between revisions of "Team:Heidelberg/Modeling/rtsms"
| Line 32: | Line 32: | ||
<div class="col-lg-12"> | <div class="col-lg-12"> | ||
<p class="basictext"> | <p class="basictext"> | ||
| + | Studying determinants of polymerase efficiency based on small-molecule sensing | ||
| + | Our subproject on small molecule sensing facilitates quantitatively studying in vitro transcription (IVT) by ATP-spinach and malachite green RNA-aptamers. In particular, we could study the inaccuracy of polymerases reflected by an excess of consumed ATP molecules over the number of ATP molecules in synthesized malachite green aptamers [1], based on a mathematical model (Fig. 3A). | ||
| + | |||
| + | Figure 4. IVT inaccuracy depends on the ATP to active template ratio. (A) A basic model with constant numbers of nA and synthesis parameters ksyn,M, was extended to variants with nA and ksyn,M depending on the polymerase concentration (Variant 2), A- and T*-dependent nA with exponents k and l (Variant 3) or only an exponent for T* (Variant 4). Fitting improvement is indicated by decreasing Akaike information criterion (AIC) values. (B) Reducing the optimal variant 4 by assuming a steady state for T*, no degradation of P or no degradation of A disproved model fits. (C) Model variant 4 can explain increasing inefficiency (higher nA) with decreasing A/T* ratios. | ||
| + | |||
<p> | <p> | ||
<img src="https://static.igem.org/mediawiki/2015/9/92/Modeling_fig4.png" style="width:100%;"> | <img src="https://static.igem.org/mediawiki/2015/9/92/Modeling_fig4.png" style="width:100%;"> | ||
| − | + | Figure 3. IVT model reactions and fits to experimental data. (A) Model reactions describing reversible assembly of templates T and polymerase P to active templates T* that incorporate ATP A into malachite green RNA-aptamers M but also into abortion products, leading to a higher number nA of consumed than ATP molecules nA,M incorporated in malachite green aptamers. (B) Model fits to data at two different polymerase concentrations. | |
| + | Because of two unexpected findings, a basic model could not explain the data. Counter-intuitively, malachite green showed a linear increase while the ATP-spinach intensity was exponentially decreasing. Furthermore, it was surprising that doubling the amount of polymerase increased the production of malachite green by even more than two-fold. Both phenomena could be explained by an optimal model variant (Fig. 3B, 4A), in which the polymerase inaccuracy increased with increasing ratios between ATP and active templates that could not be further reduced without losing fit quality (Fig. 4B, 4C). | ||
</p> | </p> | ||
| − | + | ||
| + | Figure 3. IVT model reactions and fits to experimental data. (A) Model reactions describing reversible assembly of templates T and polymerase P to active templates T* that incorporate ATP A into malachite green RNA-aptamers M but also into abortion products, leading to a higher number nA of consumed than ATP molecules nA,M incorporated in malachite green aptamers. (B) Model fits to data at two different polymerase concentrations. | ||
| + | Because of two unexpected findings, a basic model could not explain the data. Counter-intuitively, malachite green showed a linear increase while the ATP-spinach intensity was exponentially decreasing. Furthermore, it was surprising that doubling the amount of polymerase increased the production of malachite green by even more than two-fold. Both phenomena could be explained by an optimal model variant (Fig. 3B, 4A), in which the polymerase inaccuracy increased with increasing ratios between ATP and active templates that could not be further reduced without losing fit quality (Fig. 4B, 4C). | ||
<p> | <p> | ||
</p> | </p> | ||
Revision as of 04:47, 2 October 2015

Studying determinants of polymerase efficiency based on an aptamer sensor
Studying determinants of polymerase efficiency based on small-molecule sensing Our subproject on small molecule sensing facilitates quantitatively studying in vitro transcription (IVT) by ATP-spinach and malachite green RNA-aptamers. In particular, we could study the inaccuracy of polymerases reflected by an excess of consumed ATP molecules over the number of ATP molecules in synthesized malachite green aptamers [1], based on a mathematical model (Fig. 3A). Figure 4. IVT inaccuracy depends on the ATP to active template ratio. (A) A basic model with constant numbers of nA and synthesis parameters ksyn,M, was extended to variants with nA and ksyn,M depending on the polymerase concentration (Variant 2), A- and T*-dependent nA with exponents k and l (Variant 3) or only an exponent for T* (Variant 4). Fitting improvement is indicated by decreasing Akaike information criterion (AIC) values. (B) Reducing the optimal variant 4 by assuming a steady state for T*, no degradation of P or no degradation of A disproved model fits. (C) Model variant 4 can explain increasing inefficiency (higher nA) with decreasing A/T* ratios.
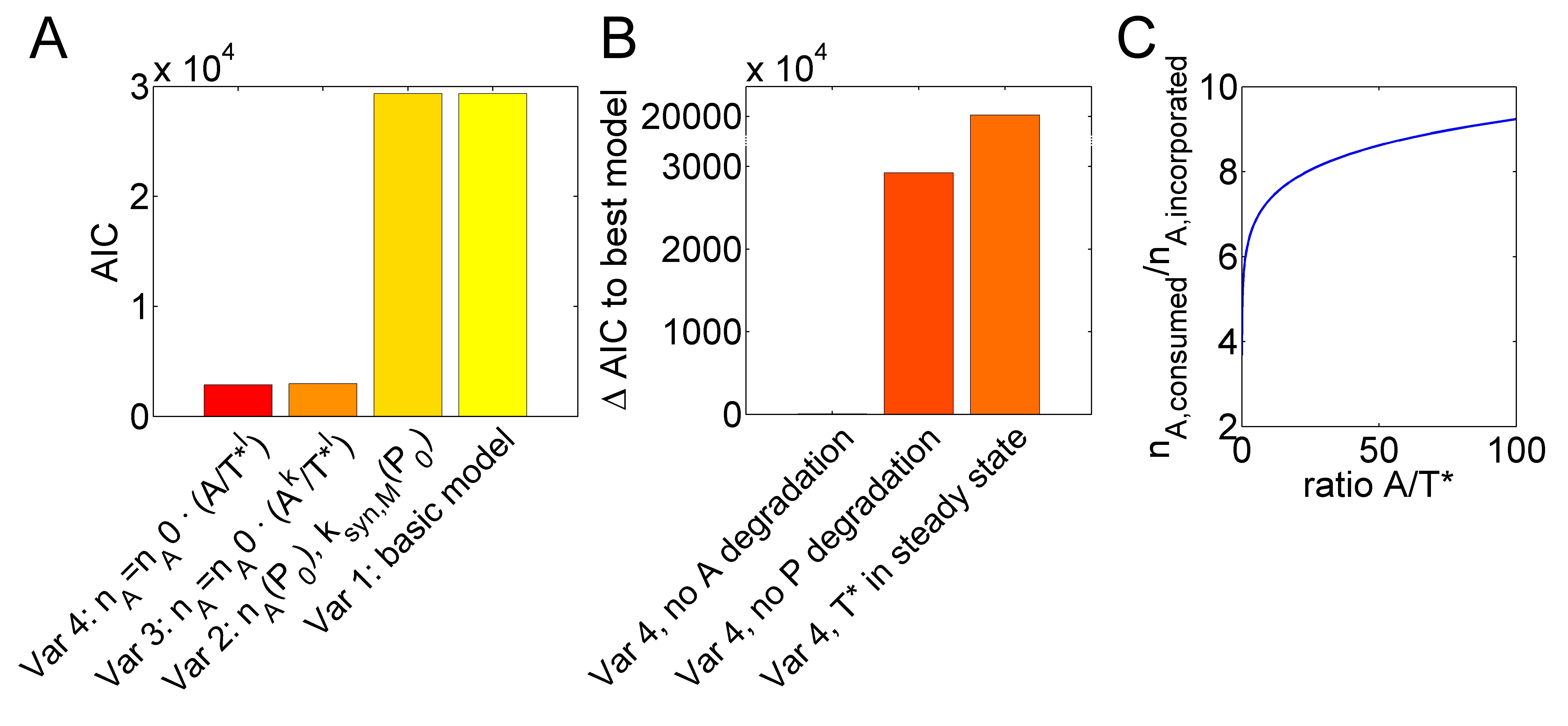 Figure 3. IVT model reactions and fits to experimental data. (A) Model reactions describing reversible assembly of templates T and polymerase P to active templates T* that incorporate ATP A into malachite green RNA-aptamers M but also into abortion products, leading to a higher number nA of consumed than ATP molecules nA,M incorporated in malachite green aptamers. (B) Model fits to data at two different polymerase concentrations.
Because of two unexpected findings, a basic model could not explain the data. Counter-intuitively, malachite green showed a linear increase while the ATP-spinach intensity was exponentially decreasing. Furthermore, it was surprising that doubling the amount of polymerase increased the production of malachite green by even more than two-fold. Both phenomena could be explained by an optimal model variant (Fig. 3B, 4A), in which the polymerase inaccuracy increased with increasing ratios between ATP and active templates that could not be further reduced without losing fit quality (Fig. 4B, 4C).
Figure 3. IVT model reactions and fits to experimental data. (A) Model reactions describing reversible assembly of templates T and polymerase P to active templates T* that incorporate ATP A into malachite green RNA-aptamers M but also into abortion products, leading to a higher number nA of consumed than ATP molecules nA,M incorporated in malachite green aptamers. (B) Model fits to data at two different polymerase concentrations.
Because of two unexpected findings, a basic model could not explain the data. Counter-intuitively, malachite green showed a linear increase while the ATP-spinach intensity was exponentially decreasing. Furthermore, it was surprising that doubling the amount of polymerase increased the production of malachite green by even more than two-fold. Both phenomena could be explained by an optimal model variant (Fig. 3B, 4A), in which the polymerase inaccuracy increased with increasing ratios between ATP and active templates that could not be further reduced without losing fit quality (Fig. 4B, 4C).
Table 3. Stepwise changes from the basic model to the optimal variant 4 and to simplifications of variant 4 to variants 4a to 4c
|
Model variant |
Subsequent modifications relative to basic model or previous variant |
Changes in fitting quality |
|
1 |
Michaelis-Menten instead of linear kinetics for active template |
no improvement |
|
2 |
Individual $k_{syn}$ and $n_A$ values for different polymerase concentrations |
improvement |
|
3 |
$n_A$ depends on function of $T_{act}$ and $A$ $n_A=n_{A,0} A^{k} /T_{act}^{l}$ |
improvement, $k\approx0$
|
|
4, best model |
Setting $k=0$ |
improvement |
|
4a |
No degradation of P in variant 4 |
decrease |
|
4b |
No degradation of A in variant 4 |
decrease |
|
4c |
Binding of $P$ to $T$ in steady state in variant 4 |
decrease |
Table 4. Model equations for the basic model and Variants 1 to 4c
|
Model species |
Variant |
Equation |
|
$P$ |
Basic model Variants 1 to 4, 4c |
$\frac{d[P]}{dt}=-k_{on}[T][P]+k_{off}[T_{act}]-k_{deg,P}[P]$ |
|
Variant 4a |
$[P](t)=[P](t_{0})\exp\left(-k_{deg,P}t\right)$ |
|
|
Variant 4b |
$\frac{d[P]}{dt}=-k_{on}[T][P]+k_{off}[T_{act}]$ |
|
|
$T$ |
Basic model Variants 1 to 4, 4b, 4c |
$\frac{d[T]}{dt}=-k_{on}[T][P]+k_{off}[T_{act}]$ |
|
Variant 4a |
$[T]=[T_{tot}]-[T_{act}]$ |
|
|
$T_{act}$ |
Basic model Variants 1 to 4, 4b, 4c |
$\frac{d[T_{act}]}{dt}=k_{on}[T][P]-k_{off}[T_{act}]$ |
|
Variant 4a |
$[T_{act}]=\frac{[T_{tot}][P]}{K_{d,P}}$ |
|
|
$A$ |
Basic model Variants 2 to 4, 4a, 4b |
$\frac{d[A]}{dt}=-k_{syn}[A][T_{act}]-k_{deg,A}[A]$ |
|
Variant 1 |
$\frac{d[A]}{dt}=-k_{syn}\frac{[A][T_{act}]}{K_{m,T}+[T_{act}]}-k_{deg,A}[A]$
|
|
|
Variant 4c |
$\frac{d[A]}{dt}=-k_{syn}[A][T_{act}]$ |
|
|
$M$ |
Basic model, Variant 2 |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A}}[A][T_{act}]$ |
|
Variants 1 |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A}}\frac{[A][T_{act}]}{K_{m,T}+[T_{act}]}$ |
|
|
Variant 3 |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A,0}\frac{[A]^{k}}{[T_{act}]^{l}}}[A][T_{act}]=\frac{k_{syn}}{n_{A,0}}[A]^{1-k}[T_{act}]^{1+j}$ |
|
|
Variants 4, 4a, 4b, 4c |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A,0}\frac{[A]}{[T_{act}]^{l}}}[A][T_{act}]=\frac{k_{syn}}{n_{A,0}}[T_{act}]^{1+j}$ |