Team:Heidelberg/Modeling/rtsms

Studying determinants of polymerase efficiency based on an aptamer sensor
Our subproject on small molecule sensing facilitates quantitatively studying in vitro transcription (IVT) by ATP-spinach and malachite green RNA-aptamers. In particular, we could study the inaccuracy of polymerases reflected by an excess of consumed ATP molecules over the number of ATP molecules in synthesized malachite green aptamers, based on a mathematical model (Figure 1A).
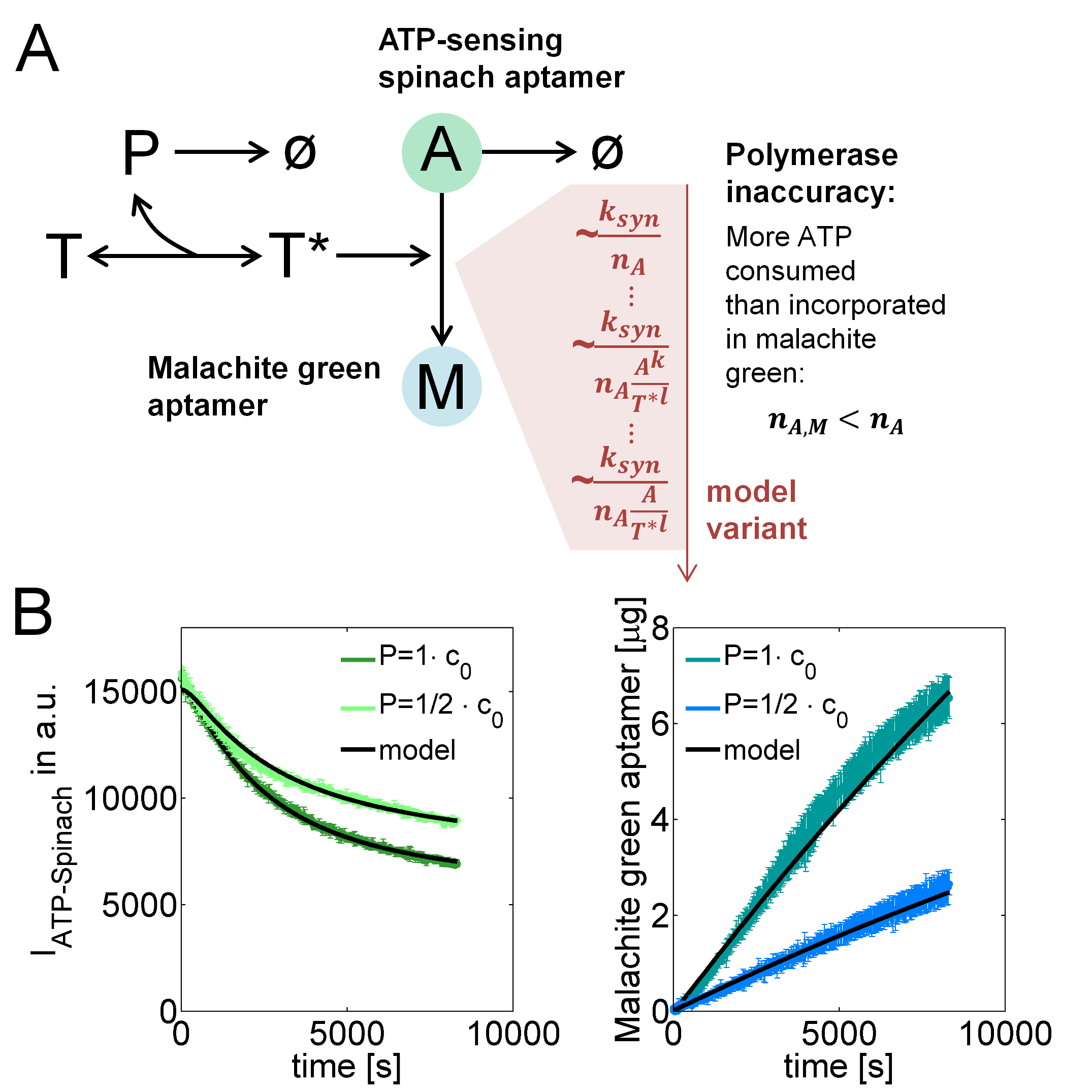
Figure 1. IVT model reactions and fits to experimental data. (A) Model reactions describing reversible assembly of templates $T$ and polymerase $P$ to active templates $T^*$ that incorporate ATP $A$ into malachite green RNA-aptamers $M$ but also into abortion products, leading to a higher number $n_A$ of consumed than ATP molecules $n_{A,M}$ incorporated in malachite green aptamers. (B) Model fits to data at two different polymerase concentrations.
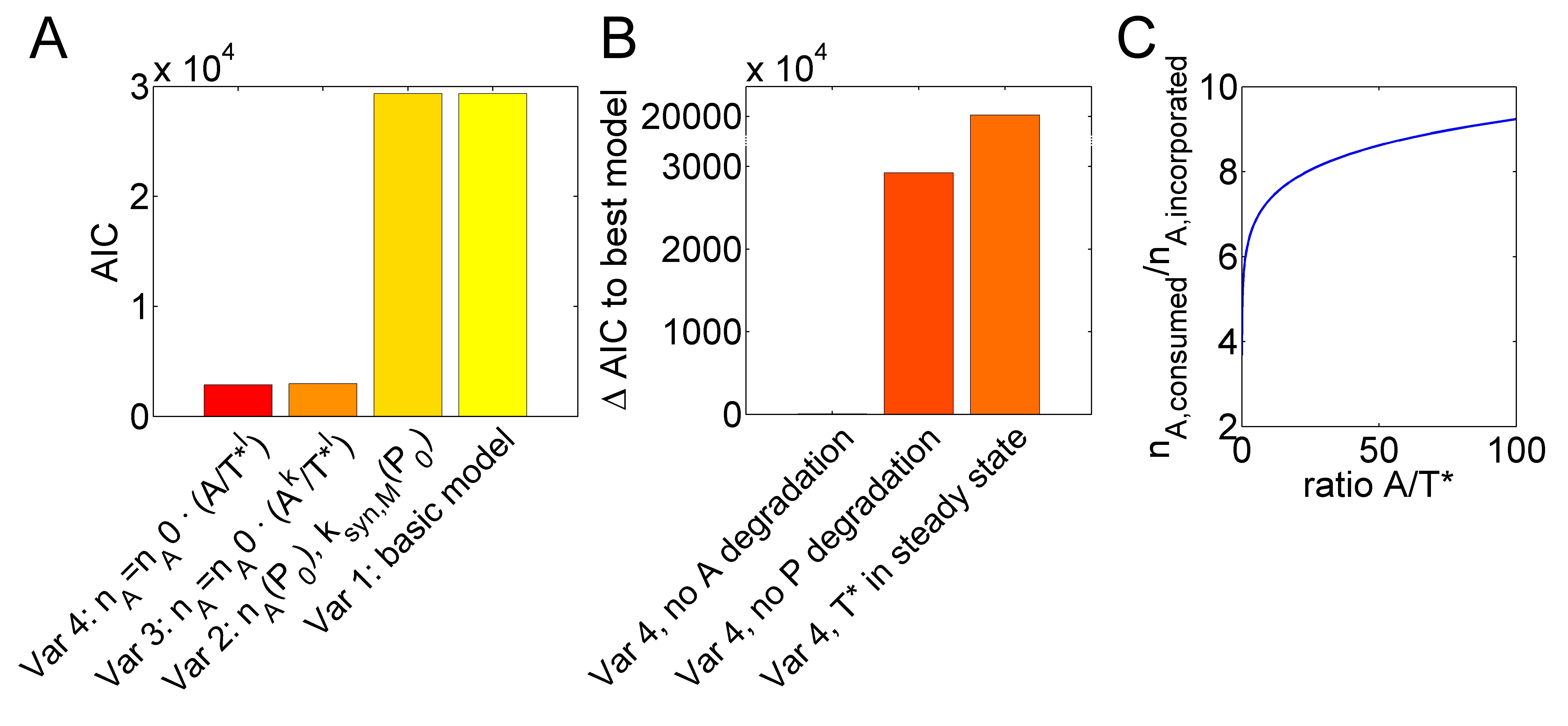
Figure 2. IVT inaccuracy depends on the ATP to active template ratio. (A) A basic model with constant numbers of $n_A$ and synthesis parameters $k_{syn,M}$, was extended to variants with $n_A$ and $k_{syn,M}$ depending on the polymerase concentration (variant 2), $A$- and $T^*$-dependent $n_A$ with exponents $k$ and $l$ (variant 3) or only an exponent for $T^*$ (variant 4). Fitting improvement is indicated by decreasing Akaike information criterion (AIC) values. (B) Reducing the optimal variant 4 by assuming a steady state for $T^*$, no degradation of $P$ or no degradation of $A$ disproved model fits. (C) Model variant 4 can explain increasing inefficiency (higher $n_A$) with decreasing $A/T^*$ ratios.
Table 3. Stepwise changes from the basic model to the optimal variant 4 and to simplifications of variant 4 to variants 4a to 4c
|
Model variant |
Subsequent modifications relative to basic model or previous variant |
Changes in fitting quality |
|
1 |
Michaelis-Menten instead of linear kinetics for active template |
no improvement |
|
2 |
Individual $k_{syn}$ and $n_A$ values for different polymerase concentrations |
improvement |
|
3 |
$n_A$ depends on function of $T^*$ and $A$ $n_A=n_{A,0} A^{k} /T^{*l}$ |
improvement, $k\approx0$
|
|
4, best model |
Setting $k=0$ |
improvement |
|
4a |
No degradation of P in variant 4 |
decrease |
|
4b |
No degradation of A in variant 4 |
decrease |
|
4c |
Binding of $P$ to $T$ in steady state in variant 4 |
decrease |
Table 4. Model equations for the basic model and variants 1 to 4c
|
Model species |
Variant |
Equation |
|
$P$ |
Basic model Variants 1 to 4, 4c |
$\frac{d[P]}{dt}=-k_{on}[T][P]+k_{off}[T^*]-k_{deg,P}[P]$ |
|
Variant 4a |
$[P](t)=[P](t_{0})\exp\left(-k_{deg,P}t\right)$ |
|
|
Variant 4b |
$\frac{d[P]}{dt}=-k_{on}[T][P]+k_{off}[T^*]$ |
|
|
$T$ |
Basic model Variants 1 to 4, 4b, 4c |
$\frac{d[T]}{dt}=-k_{on}[T][P]+k_{off}[T^*]$ |
|
Variant 4a |
$[T]=[T_{tot}]-[T^*]$ |
|
|
$T^*$ |
Basic model Variants 1 to 4, 4b, 4c |
$\frac{d[T^*]}{dt}=k_{on}[T][P]-k_{off}[T^*]$ |
|
Variant 4a |
$[T^*]=\frac{[T_{tot}][P]}{K_{d,P}}$ |
|
|
$A$ |
Basic model Variants 2 to 4, 4a, 4b |
$\frac{d[A]}{dt}=-k_{syn}[A][T^*]-k_{deg,A}[A]$ |
|
Variant 1 |
$\frac{d[A]}{dt}=-k_{syn}\frac{[A][T^*]}{K_{m,T}+[T^*]}-k_{deg,A}[A]$
|
|
|
Variant 4c |
$\frac{d[A]}{dt}=-k_{syn}[A][^*]$ |
|
|
$M$ |
Basic model, Variant 2 |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A}}[A][T^*]$ |
|
Variants 1 |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A}}\frac{[A][T^*]}{K_{m,T}+[T^*]}$ |
|
|
Variant 3 |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A,0}\frac{[A]^{k}}{[T^*]^{l}}}[A][T^*]=\frac{k_{syn}}{n_{A,0}}[A]^{1-k}[T^*]^{1+j}$ |
|
|
Variants 4, 4a, 4b, 4c |
$\frac{d[M]}{dt}=\frac{k_{syn}}{n_{A,0}\frac{[A]}{[T*]^{l}}}[A][T^*]=\frac{k_{syn}}{n_{A,0}}[T^*]^{1+j}$ |