Team:TJU/Results


Results
1 Lactate producing system
In order to verify our ldhE part and pflB knockout strategy have works well, we get following results.

Figure 1. a: The growth curve in anaerobic condition (M9 medium). b: Glucose consumption curve in anaerobic condition (M9 medium).
![]()
Shown as figure 1a, under anaerobic conditions, wild-type MG1655 has the same growth rate as MG1655ΔpflB, but MG1655ΔpflB + ldhE keeps relatively lower growth rate than the two strains mentioned before. Shown as figure 1b, the anaerobic glucose consumption rate keeps the nearly same among MG1655, MG1655ΔpflB and MG1655ΔpflB + ldhE.
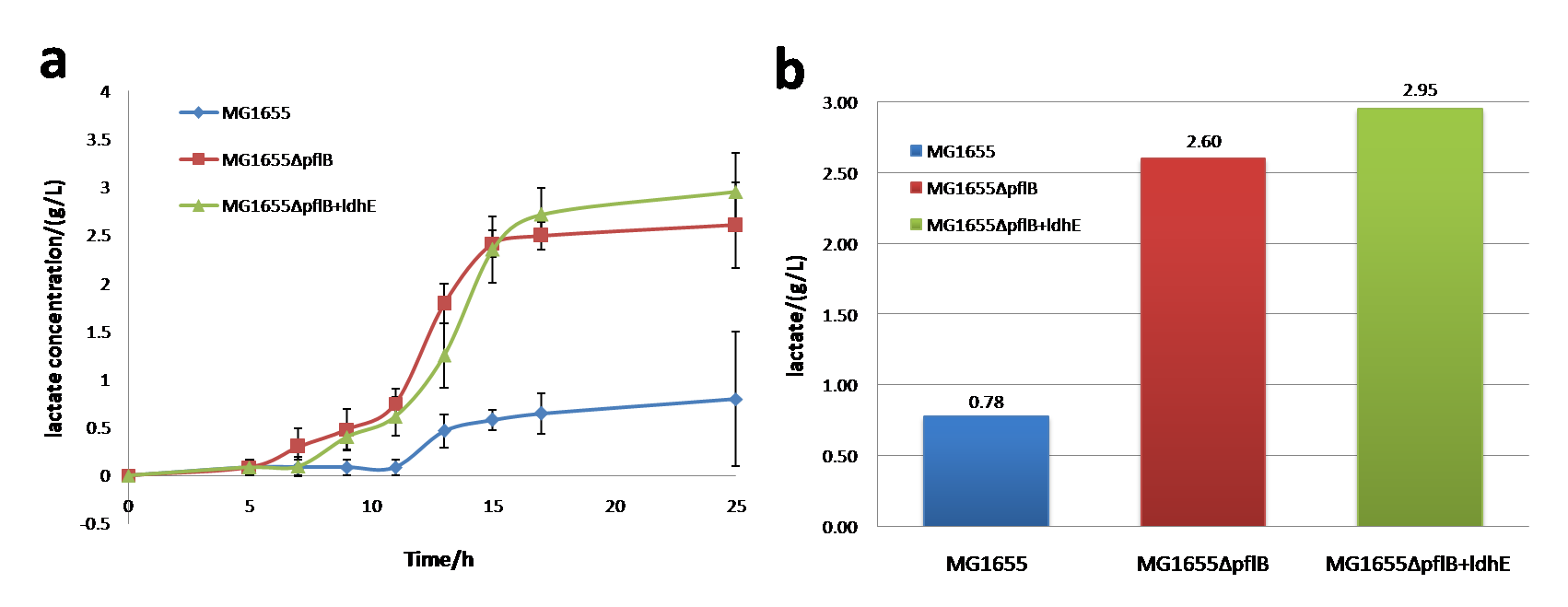
Figure 2. a: The lactate production curve in anaerobic condition (M9 medium). b: The lactate production in anaerobic condition (M9 medium, 25h after inoculation)
![]()
In anaerobic environment, lactate production of MG1655, MG1655ΔpflB and MG1655ΔpflB+ldhE has large differences between each other. Notably, knocking out of pflB together with ldhE insertion will increase lactate generation up to ~3 g/L and the amount produced by MG1655ΔpflB is smaller than MG1655ΔpflB+ldhE.
So, knocking out of pflB can dramatically improve the lactate production. Besides, ldhE also plays an important role in lactate increase. Engineered lactate producing strain (MG1655ΔpflB+ldhE) has a greater utilization of carbon sources and a higher yield of lactate with M9 medium under the same concentrations of glucose.
2 Flavins producing system
Shewanella oneidensis MR-1, a facultative anaerobe, has been widely used as a model anode biocatalyst in microbial fuel cells (MFCs) due to its easiness of cultivation, adaptability to aerobic and anaerobic environment and both respiratory and electron transfer versatility. [8]
Particularly, EET pathway can be subdivided into direct EET and mediated EET and the latter one limits the efficiency of electron transfer between bacteria and electrode due to the deficiency of electron mediator. To regulate relations of energy and information in the consortium, flavins hold the key to success.
It is widely accepted that interfacial EET is the rate-limiting step in the EET processes which can be relieved by some redox active molecules such as quinines and metal-centered porphyrin-ring derivatives. However, those redox active molecules are generally costly and toxic to anodic electricigens. Besides, Shewanella is capable of utilizing self-secreted flavins like riboflavin (RF) to accelerate EET, which is much more efficiently than other exogenous active molecules. More specifically, riboflavin (RF) and flavin mononucleotide (FMN) enhance EET more than five times, with much lower concentrations than those needed for anthraquinone-2,6-disulfonate (AQDS) shuttling. So we choose flavins as one entry point to enhance the electricity output of MFCs. [9]

Figure 9. Comparison of co-factor EET model(a) and diffusion EET model(b), (b) also reveals the electron pathway by endogenous flavins
![]()
In EET model, outward current flows from interior of cells to outer membrane (OM) and extracellular anodes through a metal-reducing conduit (Mtr pathway), where electrons (from NADH, the intracellular electron carrier) flow through the menaquinol pool, CymA (inner membrane [IM] tetraheme c-type cytochromes [c-Cyts]), MtrA (periplasmic decaheme c-Cyts), MtrB (β-barrel trans-OM protein) and finally to MtrC and OmcA (two OM decaheme c-Cyts). However, the principle of how flavins function still remains controversial. [10]
Traditional model demonstrates that flavins carry the electrons from OM c-Cyts to electrode by diffusion, which has been in debate due to thermodynamic disproof. Recently, another interfacial EET model was proposed, where flavin may serve as a co-factor binding to OmcA or MtrC to dictate EET. Fully oxidized flavin (Ox) accepts one electron from reduced heme of OM c-Cyts, and binds to OM c-Cyts as a cofactor in the semiquinone (Sq) form with shifted potential. Ox/Sq redox cycling in OM c-Cyts donates electrons to electrodes in a one electron reaction mode via direct contact.[10]

Figure 10. Pathway for engineered riboflavin and FMN production E. coli.
![]()
Although Shewanella uses endogenous flavins to mediate electrons, the amount of its production is deficient and multi-tasks brought by self-engineering may also reduce the transfer efficiency. As a consequence, we had two strategies to construct the zymophyte and further improve the production. Firstly, we introduced flavin producing genes using the E.coli as chassis. Secondly, we get an engineered B.subtilis strain from the lab of Dr. Tao Chen [11](more detail in attribution)
Based on the EET pathway theory, we suggest that by maximizing the amount of flavins, we may significantly enhance the EET efficiency and achieve a relatively high power generation.
We found a part BBa_K1172303 in Part Registry constructed by 2013 Team Bielefelf-Germany which was also aimed at producing riboflavins. However, the gene cluster showed a maximum output of 6 mg/L even with a strong promoter, which was insufficient to maintain a high and constant efficiency of EET pathway in our co-culture system. So we decided to optimize the part BBa_K1172303 to enhance the production of riboflavins.

Figure 11. Two different parts we designed to produce flavins.
![]()
As we learn from metabolic flux (figure 2), it reveals the relevant pathways of riboflavin production and engineering strategies for riboflavin production. In their previous research(Tao, et al) [7] ,they construct a high-yield E.coli strain with a yield of 229.1 mg/L. Based on their study, we constructed a flavin producing part (ribABDEC cluster) named EC10.(as shown in figure 11a). The part(BBa_K1696011) we designed, compared with BBa_K117230, has been well optimized and the yield of that reached 17 mg/L.( as shown in figure 13). We can see from the results, the functionality of their parts has successfully improved.

Figure 12. The production of EC10 in tubes.
![]()
In the meanwhile, based on co-factor model, EET pathway points out that FMN plays a critical role in electron transfer. Through the study by Dr. Tao Chen, they have weakened the RBS upstream of ribC to divert more of the material flux to RF production.[7] Based on their viewpoint, we got further to design a strong RBS sequence upstream of the ribC and we rename the new part as EC10*(BBa_K1696010). The yield of EC10* reached 90 mg/L(as shown in fugure 11b). The strong RBS before ribC sequence can lead to the FMN accumulation while riboflavin consumes a bit, which may in turn, result in increase both for riboflavin and FMN as a whole.

Figure 13. The yield of riboflavin in different strains :EC10, Rf02S (Δpgi+EC10), EC10*
![]()
When we characterized our part, we were not able to detect the FMN for lacking proper equipments. However, we were surprised to find that the production of EC10* was even better with a larger output of riboflavins. For riboflavin measurements, culture samples were diluted with 0.05 M NaOH to the linear range of the spectrophotometer and the A444 was immediately measured, according to the Dr. Tao Chen’s method [7]. As for the reason of production improvement of RF, we speculate that the strengthening of ribC gene can improve both the RF and FMN yield through the flavin metabolic pathway.
3 Co-culture MFC -- Labor Division
Figure 12 13
The relations between current density and voltage is referred as polarization curve while the power curve can represent the relations between output power density and current density. We can learn from the fugure ***, the highest power density of (1) reaches 10 mW/m2 and (2) reaches 17 mW/m2.
Figure 14 15
Figure 13. (a) The comparison of polarization curve and power curve among Shewanella, Shewanella + MG1655, Shewanella + ΔplfB ldhE + Rf02S.
(b)The comparison of polarization curve and power curve among Shewanella, Shewanella + MG1655 and Shewanella + ΔplfB ldhE + B. Subtilis.
![]()
It is obvious that the power output in three-strain MFC systems with a more complete labor division are far greater than single-strain and two-strian MFCs.
Figure 16
Figure 14. The comparison of polarization curve and power curve among Shewanella + ΔplfB ldhE + B. Subtilis and Shewanella + ΔplfB ldhE + Rf02S.
![]()
It has been shown in power curve that Shewanella + ΔplfB ldhE + B. Subtilis has a significantly higher output than Shewanella + ΔplfB ldhE + Rf02S, which indicates that three-species system can generate higher electricity and have a better MFC performance, in return, have a promising application.
E-mail: ggjyliuyue@gmail.com |Address: Building No.20, No.92 Weijin road, Tianjin University, China | Zip-cod: 300072
Copyright 2015@TJU iGEM Team